|
|

|
Plausible Mechanisms for 70-Million-Year-Old T. rex DNA Preservation and Sequenc
| .,,.,.,,.,..,.,.,.. | 05/23/25 |
 Poast new message in this thread
Poast new message in this thread

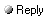
Date: May 23rd, 2025 11:44 PM Author: .,,.,.,,.,..,.,.,.. ing Introduction After 70 million years, the odds of recovering intact Tyrannosaurus rex DNA are vanishingly low. DNA is an unstable molecule that breaks down over time due to hydrolysis, oxidation, and background radiation. Under normal conditions, even in permafrost, DNA’s chemical bonds decay to mere fragments after a few million years. In fact, studies estimate DNA has a half-life of only a few hundred years, meaning after about 6–7 million years at cool temperatures most nucleotide bonds would be broken. The current record for the oldest authentic DNA comes from permafrost-preserved samples on the order of 1–2 million years old – orders of magnitude younger than the late Cretaceous era. Given these challenges, no credible sequence from non-avian dinosaur DNA has been obtained to date. Yet, to explore scientifically plausible (if extremely unlikely) scenarios for T. rex DNA survival, we must imagine exceptional conditions and emerging technologies. Below we outline several scenarios in which ~70-million-year-old T. rex DNA might survive far beyond the usual limit and how future biotechnology might recover and sequence it. Each scenario is evaluated for its mechanism of preservation, required technological interventions, and feasibility, with an emphasis on real-world science rather than science fiction. Challenges of DNA Survival Over Deep Time Before diving into scenarios, it’s important to grasp why DNA normally disintegrates long before 70 million years: Chemical Decay: DNA depurination (loss of bases) and strand breakage occur even at low temperatures. Experiments modeling DNA decay in bone suggest that even at −5 °C, DNA fragments would average only 1 base pair after ~6.8 million years. Warmer conditions accelerate this decay. Microbial Degradation: In a carcass or buried bone, bacteria and fungi rapidly consume organic molecules, including DNA, unless the remains are sealed from microbes. Radiation: Natural background radiation (from cosmic rays or radioactive isotopes in soil) causes cumulative DNA damage over geological timescales. Without exceptional shielding, this radiation would induce strand breaks and base damage that accumulate over millions of years. Known Limits: The upper bound for recoverable DNA under best conditions (cold, dry, stable) is estimated around 1–2 million years. Indeed, DNA from a ~1.65-million-year-old mammoth and environmental DNA ~2 million years old in Greenland sediments have been sequenced. Beyond this, even trace DNA becomes vanishingly scarce and exceedingly fragmented. Given these challenges, any scenario for 70-million-year survival requires extraordinary preservation conditions that slow or halt the usual decay processes, coupled with cutting-edge retrieval methods to piece together extremely damaged genetic material. Below we consider several such scenarios. Scenario 1: Resin Entombment (Amber) of Tyrannosaur DNA One iconic idea for preserving dinosaur DNA is entombment in tree resin, which fossilizes into amber. In theory, amber can encase biological material in a protective tomb, sealing out oxygen and microbes. For T. rex, the plausible route is indirect: for example, a blood-sucking insect (tick or mosquito) might have fed on a T. rex and then become trapped in resin, preserving traces of the dinosaur’s blood (and DNA) in its gut. Amber preservation can be astonishingly detailed – it can protect actual cells and soft tissues (even stomach contents) of organisms across tens of millions of years. This inspired the famous Jurassic Park concept of DNA retrieval from dino-blood in insects.
A fossil insect (stingless bee in ~Copal amber) encased in resin. Amber entombment preserves fine details of organisms, but DNA survival in amber over tens of millions of years has proven very elusive. Why it Could Work: Resin hardens and isolates inclusions from water and air, potentially creating a microenvironment where DNA decay is slowed. Without water, hydrolytic reactions that break DNA are limited. Amber also blocks UV radiation. In the mid-1990s, some scientists claimed to have extracted DNA from Cretaceous amber inclusions – for example, a team reported ~120–130 million-year-old DNA fragments from an amber-trapped weevil latimes.com. This was 80 million years older than any previously known DNA at the time, suggesting amber as a “molecular time capsule.” They recovered only a tiny amount (on the order of 1 millionth of the genome) – not even a full gene – but it hinted that bits of DNA might survive in amber. The scientists were optimistic that amber could be a “treasure chest for molecular paleontologists.” Indeed, they immediately began searching for dinosaur blood DNA in a 75-million-year-old amber midge. Challenges and Feasibility: Unfortunately, later, more rigorous research has cast serious doubt on amber as a DNA reservoir. A 2013 study attempted to extract DNA from insects in copal (sub-fossilized resin, only 10,000 years to 1 million years old) under sterile, contamination-free conditions. They found no authentic ancient DNA at all, even in these much younger resin specimens. This suggests that resin entombment does not effectively halt DNA decay; DNA in amber may degrade as fast as in dry air. The supposed Mesozoic DNA results from the 1990s are now thought likely to be contamination or false positives (e.g. modern DNA amplified by mistake). One intuitive might think “instantaneous” resin burial would preserve DNA, but empirical evidence indicates otherwise. For T. rex specifically, amber entombment would be an even rarer scenario – large animals aren’t directly caught in amber. It would rely on an intermediary (parasite or perhaps plant material with T. rex saliva, etc.). Even if such an inclusion were found, any surviving DNA would be in tiny fragments mixed with insect DNA. Recovering it would require ultra-sensitive sequencing. Current next-generation sequencing (NGS) can read all DNA in a sample without PCR bias, so theoretically one could shotgun-sequence an amber insect’s gut contents and look for tell-tale dinosaur sequences. We’d need to distinguish them from microbes and modern contamination by characteristic damage patterns (e.g. C→T substitutions from ancient cytosine deamination) and by similarity to bird/reptile DNA rather than to modern species. Targeted enrichment could also help – e.g. using probes based on avian DNA to capture fragments that might be dinosaurian. Assessment: Real-world plausibility is extremely low. The protective effect of amber appears limited – DNA likely degrades too much over tens of millions of years. We have yet to reproducibly detect even Cenozoic insect DNA in amber. Thus, while resin entombment is a classic scenario, it must be viewed skeptically. It would require an almost miraculous convergence of factors (a T. rex blood meal in amber, DNA fragments surviving despite chemical and radiative damage, and next-gen sequencing powerful enough to fish out a few dozen authentic base pairs). Even under near-future tech, this scenario remains highly speculative and would likely yield at best minuscule fragments if any. As one amber DNA expert concluded: “unfortunately, the Jurassic Park scenario must remain in the realms of fiction.”
Scenario 2: Cryogenic Preservation in Permafrost or Ice Extreme cold is known to slow chemical reactions and biological activity, making it the friendliest environment for long-term DNA survival. One could imagine a scenario in which T. rex tissue was flash-frozen and remained at subzero temperatures for eons. For example, if a Tyrannosaur died in a high-latitude environment that later became permafrost, its remains might stay partly frozen from that point on. While the late Cretaceous climate was generally warm, there were polar dinosaurs in regions like Alaska and Siberia. It’s hypothesized that some tyrannosaurid (or a T. rex carcass transported by rivers) could end up buried in sediments that eventually froze during Cenozoic cooling. If the remains stayed continuously frozen (and preferably dry and dark) from, say, a few million years after the dinosaur’s death until now, DNA decay could have been drastically reduced. Why it Could Work: In known cases, permafrost has preserved DNA far longer than temperate climates. The previous record for oldest genomic DNA was a ~700,000-year-old horse from Canadian permafrost, whose genome was sequenced successfully atlasobscura.com . More recently, DNA fragments 2 million years old were recovered from frozen sediments in Greenland scientificamerican.com . Cold not only slows chemical bond breakage but also inhibits microbial action (organisms go dormant or cannot thrive). If a T. rex bone and marrow, for instance, lay deeply frozen, the DNA inside cells would be largely shielded from liquid water and microbial enzymes for millions of years. In principle, at sufficiently low temperature (e.g. deep-frozen at < −10 °C continuously), DNA’s half-life can extend significantly, meaning some fragments might persist far longer than in a warm setting en.wikipedia.org . Challenges and Feasibility: The obvious issue is that continuous 70-million-year freeze is geologically implausible. There was no permafrost in the Cretaceous – polar regions were much warmer. The earliest deep freezes (in high latitudes) occurred in the late Cenozoic (within last few million years). This means a T. rex carcass would have had to somehow “time travel” through tens of millions of years of warmer conditions before entering a permafrost state. Perhaps one could contrive that the DNA was preserved by other means initially and then locked in permafrost during the Quaternary glaciations – a combo scenario. However, absent magical timing, a true cryogenic preservation from day one is unrealistic. Even organisms that did get frozen in the Mesozoic (if any) would have experienced thaw cycles, geothermal heat, etc., over such vast timescales. Even assuming a miraculous unbroken freeze, radiation still works at cryogenic temperatures. Over 70 million years, natural background radiation (from e.g. uranium traces in soil or cosmic rays) would cause cumulative damage. Only heavy shielding (thick ice sheets or rock) could attenuate this, and even then not fully. For context, 1–2 million-year DNA is our upper bound from permafrost; 70 Myr is >30 times older. Extrapolating decay kinetics suggests even in ice, the DNA would fragment to mere oligonucleotides (5–10 bp) or chemical residues over that timeframe en.wikipedia.org . From a technology standpoint, if any T. rex DNA did remain in ice, it would be in extremely poor condition and ultra-low quantity. We would apply the full arsenal of ancient-DNA methods: purification in clean rooms, enzymatic damage repair (e.g. treating extracts with DNA repair enzymes to fix nicks and oxidative lesions), and single-stranded DNA library prep to capture short fragments. High-throughput sequencing would then read millions of sequences; bioinformatics could sift for those bearing the hallmark ancient damage pattern (e.g. overrepresentation of C→T changes at fragment ends) and matching phylogenetically near bird/reptile sequences. We might target high-copy loci like mitochondrial DNA (hundreds of copies per cell) to increase odds of recovery. If a contiguous freeze really preserved some fragments, we could hope to retrieve short mitochondrial or nuclear gene segments. Assessment: This scenario’s plausibility is extremely limited by geology – continuous deep freeze for tens of millions of years has not occurred on Earth. It is more of a thought experiment in pushing the known limits. However, it’s scientifically grounded in that cold is demonstrably the best preservative for DNA, so if one were to “cheat” the usual time limit, extreme cryostasis is a logical necessity. If somehow a T. rex sample were frozen in a polar environment and then never thawed, it is conceivable that traceable DNA fragments (perhaps a few tens of base pairs long) might survive. With expected near-future sequencing improvements – even sequencing single molecules base-by-base – we might then detect and read those traces. Realistically though, without some additional preservation factor, 70 million years stretches credulity; this scenario is only slightly less fanciful than amber, and would likely yield only partial minuscule sequences at best. Scenario 3: Mineral Entrapment in Fossil Bone and Crystals Fossilization can sometimes be a friend to biomolecular preservation. If DNA becomes bound to or encapsulated by minerals early in the fossilization process, it might be shielded from degradation. Bone itself is a mineral-organic matrix (mostly hydroxyapatite crystals). In younger fossils (thousands to a few hundred thousand years), it’s known that DNA often survives longest when adsorbed to bone’s mineral phase or hidden in dense bone regions. We propose that in exceptional cases, a T. rex’s bones or other tissues could have undergone fossilization in a way that locked in tiny pockets of original DNA, protecting them from water, oxygen, and microbes. Mechanism: Rapid burial in the right sediment can promote early diagenetic mineralization. For example, silica-rich water could percolate the bones and precipitate microscopic silica (opal) or calcite crystals, entombing organic molecules within. If small fragments of DNA were trapped in such mineral microspheres or crusts, they’d be less exposed to chemical attack. Similarly, DNA can adsorb to clays or sands – a known phenomenon even in soils where clay binding extends DNA persistence by shielding from nucleases sciencedirect.com onlinelibrary.wiley.com . A fossil bone might act like a molecular trap: as it mineralized, some of the original cellular material (including nuclei or collagen fibers) could get coated with minerals, creating a sealed microenvironment. Crucially, if groundwater is kept out (perhaps by a tight mineral concretion), the DNA inside might avoid complete hydrolysis. There is tantalizing evidence that original biomolecules can survive in dino bones. Paleontologist Mary Schweitzer famously found flexible blood vessel structures and protein fragments (collagen) in a 68–80 million-year-old T. rex and hadrosaur bones news.ncsu.edu news.ncsu.edu . These vessels were protected within dense bone, and analyses confirmed they contained dinosaurian collagen protein, not microbial biofilm nature.com nature.com . Collagen is more robust than DNA, but its survival suggests an unusually preserving microenvironment – possibly aided by iron (we address that in Scenario 4) and by being mineral-cradled. More directly, recent studies reported that dinosaur cartilage cells from a 75-million-year-old Hypacrosaurus (duck-billed dino) fossil show structures consistent with nuclei and even chemical markers of DNA news.ncsu.edu news.ncsu.edu . Stains that bind DNA (DAPI, propidium iodide) showed localized binding in these fossil cells, whereas control stains for bacteria were negative news.ncsu.edu news.ncsu.edu . This suggests that extremely fragmentary DNA (or DNA breakdown products) might still reside within those mineralized cartilage cells.
Fossilized cartilage cells from a 75 Myr Hypacrosaurus (top) showing positive DNA stain in a small intracellular region (white arrows), compared to modern emu cartilage cells (bottom, black arrows). The faint staining in the dino cells suggests some original DNA or its chemical remnants persist, protected by calcified cartilage sci.news sci.news . Why it Could Work: Mineral binding can stabilize biomolecules. In one experiment, ostrich eggshell proteins ~3.8 million years old were successfully sequenced, far older than any DNA, because the proteins were tightly bound in the eggshell’s mineral matrix atlasobscura.com atlasobscura.com . The calcite crystals essentially “fixed” the peptides in place, reducing their degradation. By analogy, if T. rex DNA (likely already cut into short pieces after death) adsorbed to bone apatite or was encapsulated by early mineral deposits, it might have a longer effective half-life. Also, deep burial can reduce radiation (thick sediments attenuate cosmic rays) and keep temperature stable. Some extreme claims even suggest microbes or their DNA survived inside salt crystals for tens of millions of years en.wikipedia.org , though contamination doubts remain en.wikipedia.org . Still, halite (rock salt) can trap organic inclusions; one could envision, for instance, a T. rex’s blood or tissue fluid being caught in evaporitic minerals – an unusual but not impossible scenario if the animal died near a salt flat or coastal lagoon that dried out. Recovery with Technology: If DNA is mineral-bound, standard ancient DNA extraction (which often involves EDTA, a chemical to dissolve apatite) could free those fragments. The challenge is that such DNA will be ultra-short and damaged. New library preparation methods exist that can capture very short fragments (20–30 bases or even less) by using single-stranded DNA techniques and special adapters. We also use repair enzymes: for example, T4 DNA polymerase can fill in or trim overhangs caused by damage, and uracil-DNA glycosylase (UDG) can remove uracil from cytosine deamination to make the strands more readable spaam-community.org spaam-community.org . After extraction, massive sequencing depth would be needed to fish out a signal. Computationally, we’d align any recovered sequences to genomes of living relatives (birds, crocodilians) to see if they cluster phylogenetically outside modern species – a hint they derive from an ancient lineage. If multiple overlapping short reads matching the same dinosaurian genome regions are found, we could start reconstructing partial sequences. Hybridization capture could greatly assist: e.g. designing probes from conserved genes (like mitochondrial genes or rRNA genes that change slowly) to enrich any homologous fragments from T. rex. Even if each fragment is only, say, 30 bp long, having many overlapping pieces could allow researchers to mosaic together a section of T. rex DNA by aligning to a bird reference genome. This is similar in principle to how Neanderthal DNA (often highly fragmented) was assembled using the human genome as a scaffold – albeit the dino-bird divergence (~65+ Myr) is far larger, making it a bigger puzzle. Assessment: This scenario is scientifically plausible on a small scale – it aligns with empirical evidence that some molecular remnants of dinosaurs do persist when they are mineral-bound and protected news.ncsu.edu news.ncsu.edu . However, any DNA retrieved would likely be highly incomplete. We might recover snippets of genes or maybe mitochondrial segments, but not large contiguous stretches, let alone an entire genome. Still, for the first time, real dinosaur DNA sequences could be identified. Feasibly, within the next 10–20 years, improvements in ultra-short read sequencing and data analysis could make it possible to confirm and sequence these snippets. The underlying assumption is that fossilization was gentle enough to entomb some original biomolecules before they completely decayed. Not every fossil will offer that – most don’t – but a select few (perhaps those with exceptional preservation, like deep burials in anoxic, fine-grained sediments or chemical sediments) might. Compared to amber or pure cryogenic scenarios, mineral entrapment strikes a balance of realism and scientific basis: it doesn’t violate known preservation chemistry, but it also doesn’t promise dramatic amounts of DNA. It is plausible that a very lucky T. rex fossil could one day yield a few readable DNA fragments with advanced techniques. Scenario 4: Natural Chemical Stabilization (Cross-Linking “Fixatives”) Another angle to extraordinary preservation is if the T. rex’s own biochemistry or burial chemistry effectively “fixed” the tissues after death, analogous to how a preservative might. In modern labs, we use chemicals like formaldehyde to cross-link DNA and proteins, preventing decay (e.g., in tissue samples). Perhaps nature has its own analogs. Two proposed mechanisms are: Iron-mediated preservation: When an organism dies, iron from hemoglobin and other iron-rich compounds can be released. Research by Schweitzer et al. suggested that iron can catalyze chemical reactions (Fenton chemistry) that produce free radicals, which in turn cause proteins to form cross-links nature.com nature.com . Essentially, tissues get “tanned” or fixed by their own iron. This process could stabilize structures like blood vessels and collagen for eons by making them less digestible to microbes and less soluble nature.com nature.com . The downside is that free radicals might also attack DNA, but if the DNA becomes cross-linked to proteins or other molecules, it might be somewhat shielded from total loss – locked in an inert matrix. Schweitzer’s team demonstrated that adding hemoglobin to blood vessels in vitro preserved them from decay, supporting the idea that iron has a formaldehyde-like fixative effect nature.com nature.com . This could help explain the soft vessels in T. rex bone. Maillard reactions (sugar cross-linking): As cells break down, sugars can react with amino acids and DNA to form complex brown polymers (advanced glycation end products). This is the same chemistry that causes foods to brown when heated (the Maillard reaction) and can also happen slowly at ambient temperatures. In a carcass, the combination of proteins, DNA, and sugars could yield a tangled polymeric substance – effectively “toasting” the tissues. These Maillard products are relatively inert and can survive long-term. Some scientists have proposed that the dark, insoluble residues in fossils could be such cross-linked materials preserving the outlines of soft tissue. For instance, the very old (~>200 Myr) protein traces detected in fossils atlasobscura.com might be explained by cross-linking that prevented total degradation, albeit at the cost of the original molecules being chemically altered. Why it Could Work: If a T. rex’s soft tissues were naturally fixed shortly after death by these processes, the DNA within cells might avoid immediate decay. For example, a carcass in a warm, iron-rich environment (imagine a dinosaur dying in a marsh and blood seeping into tissues) might experience rapid cross-linking. Iron-induced crosslinks could tie DNA to histone proteins or cell membranes, so it doesn’t leach away or get fully broken down. Similarly, partial desiccation in the carcass (perhaps it dried out a bit in the sun before burial) could concentrate sugars and facilitate Maillard crosslinks, essentially mummifying the remains. We know that even on shorter timescales, mummified tissues (tens of thousands of years old) sometimes retain DNA. For instance, desert mummies and peat bog bodies (thousands of years old) can yield DNA, partly because drying or tanning agents (peat tannins) preserved them. Scale that concept up: a “chemically mummified” T. rex might endure longer than a water-logged one. Challenges: The major catch is that cross-linking, while preserving tissue morphology, often destroys or masks the DNA sequence. A cross-linked DNA strand is typically not readily accessible for PCR or sequencing – it’s like having the pages of a book glued together. In fossils, the DNA might not be free-floating but rather trapped in a brown polymer mass. Current sequencing requires DNA strands to be separated and copied or read; heavy crosslinks impede this. In studies of formalin-fixed tissues (where DNA is cross-linked), scientists can recover DNA by reversing some cross-links with heat or chemicals, but highly cross-linked specimens yield very short, damaged DNA at best. For a dinosaur, the crosslinks would be far more extensive and possibly irreversible. Additionally, iron can catalyze radical damage to DNA (breaking it apart) even as it cross-links proteins – a double-edged sword. One hypothesis is that iron preserves proteins better than DNA; indeed, Schweitzer’s T. rex vessels had collagen but no sequencable DNA. So, this scenario might explain protein preservation more than DNA. Needed Technologies: To get sequence out of cross-linked, “fossilized” genetic material, we’d need advanced chemical and enzymatic tricks. One approach is to break the cross-links: for example, using chelating agents to remove iron and compounds like N-phenacylthiazolium bromide (known to break some advanced glycation product cross-links) to loosen the polymer matrix. If we manage to liberate fragments of DNA, they could then be sequenced using ancient DNA protocols. Another approach is to directly read DNA in situ despite crosslinks. Emerging methods like nanopore sequencing might help: a nanopore can sometimes thread a single molecule through a pore and detect bases even if the strand is chemically modified. If cross-links are sporadic, a nanopore might still traverse the DNA, or future nanopore variants could have enzymes that cut cross-linked chunks on-the-fly. It’s speculative, but within 10–20 years, we might see sequencing tech that is much more tolerant of DNA damage and chemical modifications than today. We should also mention computational methods: if only fragments of sequence are recovered, algorithms could compare them to likely templates. Because cross-link preservation might preferentially keep certain repetitive regions (or perhaps GC-rich regions that bind proteins strongly), we might recover unrepresentative chunks. Sophisticated de novo assembly would be difficult, but if we at least identify multiple overlapping pieces, we can guess the intermediate sequences. Also, protein sequences obtained from cross-linked fossils (e.g. dinosaur collagen peptides that have been sequenced nature.com nature.com ) could guide DNA reconstruction of gene coding regions, since we could infer likely codon sequences for those proteins by comparing to relatives. Assessment: Natural chemical stabilization is a plausible mechanism for long-term preservation of soft tissue structure and possibly extremely fragmentary DNA. It likely played a role in the preservation of cells and proteins that we know exist in some dinosaur fossils news.ncsu.edu nature.com . However, retrieving DNA sequences from such preserved tissues remains a formidable challenge. The scenario assumes that some DNA isn’t completely obliterated – just heavily modified – and that future methods can tease out the original genetic information. This is not guaranteed; the preservation might be too effective, in the sense of turning the DNA into unreadable goop (like ashes of a book – the outline is there but the text is gone). Still, if even small stretches of DNA are intact within that cross-linked matrix, improved recovery techniques could find them. Of all scenarios, this one emphasizes “stability through stasis” – the DNA is not so much protected by environment as by being chemically frozen in time. It’s a long shot, but one grounded in real chemical processes observed in laboratory and fossil contexts. Feasibility is low-to-moderate: it might allow recovery of tiny authentic DNA fragments (a few dozen base pairs) if our extraction and sequencing tools become adept at handling heavily damaged, insoluble DNA. It’s plausible enough to warrant trying on any exceptionally preserved soft-tissue fossils, using every trick in the molecular biology toolbox. Emerging Technologies for Ancient DNA Recovery Regardless of the preservation scenario, any 70-million-year-old DNA will be highly degraded and scarce. The next 10–20 years will likely see major improvements in how we detect and read such genetic trace material. Here are key technological advances that boost the prospects of sequencing ultra-ancient DNA: Ultrasensitive, Single-Molecule Sequencing: Current NGS machines already can sequence single DNA molecules (as in nanopore or PacBio sequencing) without needing billions of copies. Future refinements will increase their accuracy and ability to call modified bases. This means even if only a handful of T. rex DNA molecules survive, we could theoretically sequence them directly. Nanopore sequencing, in particular, can thread very short fragments and might identify characteristic damage patterns (like methylation or deamination) on an ancient strand. By 20 years from now, we may have quantum sequencing or improved nano-technologies that read base sequences by tunneling currents or microscopy, potentially reading fragments too damaged for current methods. DNA Damage Repair and Reconstruction: Enzymatic repair cocktails are already used on ancient DNA extracts. These include DNA polymerases that fill in broken ends, ligases that join fragments, glycosylases that remove damaged bases, and photolyases or chemical treatments to reverse crosslinks. A continued effort here could yield specialized enzymes (perhaps evolved from extremophile microbes) that can resurrect short, damaged DNA. For example, polymerases from certain bacteria can read through lesions that stall normal enzymes. There is also research into ligation of overlapping fragments – essentially trying to puzzle small pieces together in the test tube. One imaginative idea is using a bacterial system that naturally stitches small DNA pieces into a circle (some microbes take up environmental DNA and recombine it). In the lab, one could use such enzymes to take 20–30 bp fragments and ligate overlapping ones, gradually rebuilding longer segments before sequencing. While not yet available, this “ancient DNA assembly” approach could emerge as biotechnology improves. Targeted Molecule Fishing: As mentioned earlier, hybridization capture using probes from known genomes (birds, crocs) can help enrich any relic T. rex DNA. If we suspect a certain gene might be present (say, a conserved rRNA gene), we can design RNA probes to bind any complementary dino DNA in the extract, then pull those out for sequencing. This strategy greatly increases the signal-to-noise ratio. In the future, even more clever targeting might appear – e.g. CRISPR-based enrichment, where Cas proteins programmed with ancient sequence motifs cut everything except the target pieces, or microfluidic sorting that separates molecules by some signature of ancient DNA (like presence of specific damage types or binding to certain antibodies). In fact, researchers have used antibodies that bind the DNA backbone or histones to identify dinosaur DNA remnants in cells news.ncsu.edu news.ncsu.edu – one could imagine adapting that to extract those DNA-bound histones and then sequence the DNA attached. Computational Genomics and Machine Learning: If only fragmentary sequences are recovered, how do we tell if they’re truly from T. rex? Advanced computational methods will be crucial. Phylogenetic placement algorithms can take a tiny DNA read and statistically determine if it’s closer to dinosaur lineage vs. contaminant sequences. Machine learning might be used to recognize patterns of post-mortem damage to authenticate sequences as ancient. Moreover, given some genuine fragments, computational modeling can help reconstruct probable sequences by aligning them to multiple reference genomes (e.g., multiple bird genomes, since T. rex is more distantly related to any single bird). This is like solving a puzzle with many missing pieces: algorithms can fill gaps by inference, outputting a consensus sequence with uncertainty estimates. While this would not be as good as an actual strand of DNA, it could guide researchers in synthesizing the inferred DNA for further testing. Indeed, synthetic biology might become part of the toolkit – if we have enough short reads covering a gene, we could synthesize that gene and express it in a living cell to check if it produces a functional protein, cross-validating our sequence hypothesis. Proteomic and Multi-omics Integration: Though not DNA sequencing per se, analyzing preserved proteins or even sugars from the fossil can provide genetic clues. For instance, if we sequence dinosaur collagen peptides (which has been done nature.com ), we can compare them to modern species’ collagen gene sequences to deduce likely DNA coding sequences. This can narrow down what the dino DNA would look like at those gene regions. In short, a multi-pronged approach combining any DNA recovered plus protein data and ancestral genome modeling could allow a partial reconstruction of the T. rex genome, even if the DNA itself is fragmentary. All these technologies either exist in nascent form or are realistic extensions of current trends. Over two decades, it’s reasonable to expect orders-of-magnitude improvements in sequencing sensitivity and data analysis, which is exactly what would be needed to tackle such an ancient and damaged DNA sample. Feasibility Summary of Scenarios The table below summarizes the proposed scenarios, their key assumptions, required technologies, and an appraisal of feasibility: Scenario Preservation Mechanism Key Assumptions Techniques Needed Feasibility 1. Amber Entombment Resin seals tissue (insect with T.rex blood) from environment latimes.com . Amber prevents microbial decay and water damage. Dino DNA present in amber inclusion; amber significantly slows chemical decay. Ultraclean extraction; NGS of ultra-short fragments; contamination filtering sci.news . Possibly probe capture for dino-specific sequences. Extremely low. Amber’s DNA preservation proven poor sci.news . Any DNA likely <20 bp and very scarce. Success would yield only tiny fragments. 2. Cryogenic (“Icebox Rex”) Immediate freezing and continuous cold at <0 °C for millions of years. Analogous to permafrost preservation of mammoth DNA. Unbroken deep freeze from shortly after death to present (geologically unlikely). Low radiation environment. Careful sample handling in cold; enzyme repair of bases; single-strand DNA libraries; deep sequencing to find rare fragments. Very low. Requires unrealistic climate history. Even then, 70 Myr is far beyond known cold-preservation record (2 Myr) scientificamerican.com . Only very short DNA pieces would remain. 3. Mineral Entrapment DNA bound within bone minerals or encased by early diagenetic minerals (silica, calcite, clay). Microenvironment shields DNA from water and microbes. Rapid burial and mineralization before DNA completely decays. Fossil remains undisturbed by water or heat. Demineralization (EDTA) to release DNA; ancient DNA library prep for ultra-short fragments; targeted DNA capture (using bird/croc DNA probes); high-throughput sequencing. Low-Moderate. Unusual but evidence of DNA in Mesozoic cartilage exists news.ncsu.edu . Might yield partial sequences (dozens to hundreds of bp total). Not a full genome, but scientifically valuable snippets feasible. 4. Chemical Cross-linking “Natural fixation” by iron and Maillard reactions. Tissues auto-preserve via cross-linked proteins/DNA (like formalin fixation) nature.com . Preserves structure and some molecular integrity. Dense soft tissues remain with extensive cross-links rather than rotting. DNA not completely destroyed, just immobilized. Aggressive chemical treatments to break cross-links (chelation, heating); or advanced sequencing that can read cross-linked DNA; proteomic analysis to guide DNA inference. Low (for DNA). Explains exceptional protein preservation nature.com , but DNA likely very damaged. Could preserve some DNA bits in situ news.ncsu.edu . Sequencing would be extremely challenging, potentially yielding only trace sequences.
Table: Overview of proposed survival scenarios for T. rex DNA, with preservation mechanisms, assumptions, required tech, and feasibility estimates. As the table suggests, none of these scenarios are easy or likely – each stretches the limits of preservation and detection. The most promising in terms of scientific plausibility is the mineral entrapment scenario, which has some backing evidence and could synergize with advances in ancient DNA sequencing. The others (amber, deep freeze, cross-linking) rely on more exceptional circumstances or have intrinsic hurdles in recovering readable DNA. However, all scenarios share a critical point: even in the best case, we do not expect an intact dinosaur genome or anything close. The goal would be recovering enough DNA fragments to sequence and identify – perhaps a few genes or short genomic regions that we could confidently say came from T. rex. That alone would be a massive breakthrough, offering a window into dinosaur genetics. As one geneticist said in 1993, “Even a single legible dinosaur gene would be more than enough to make us happy.” latimes.com . The scenarios above, while speculative, outline how that ambitious goal might be achieved without invoking any fictional science. Conclusion In summary, pushing DNA preservation from ~1–2 million years to ~70 million years demands drastic conditions and innovative methods. We explored how a Tyrannosaurus rex’s DNA might survive – via entrapment in amber, everlasting ice, protective mineralization, or molecular fixation – and how future technologies could resurrect sequences from the resulting chemical “fossils.” Each scenario hinges on outlier circumstances: perfectly sterile amber, a freak flash-freeze, a bone that becomes a molecular time-capsule, or a built-in biochemical preservative. While these are long shots, they remain scientifically conceivable – they do not require breaking physical laws, only extending known preservation phenomena to an extreme. Crucially, emerging biotechnologies in DNA sequencing and molecular recovery will be the enabler. The next decades will likely bring DNA sequencing methods of unprecedented sensitivity, perhaps capable of reading molecules once considered too short or damaged. As techniques improve, the threshold of recoverable ancient DNA has steadily been pushed back (from tens of thousands of years to hundreds of thousands, now to million+ years scientificamerican.com ). If this trend continues, who’s to say we won’t eventually decipher some genetic code of the dinosaurs? Realistically, we should temper expectations: any T. rex DNA we manage to sequence will probably be in fragmentary form – perhaps a few dozen base pairs here, a few hundred there. But even those fragments could confirm evolutionary relationships (e.g., affirming birds as dinosaur descendants at the DNA level) and potentially reveal unique genetic traits of Tyrannosaurs. The pursuit is worthwhile, as long as it is grounded in rigorous contamination controls and analytical skepticism (many past claims of “dino DNA” have fallen apart under scrutiny en.wikipedia.org ). In the end, 70 million years is an eye-blink in geologic time but an eternity for DNA. By combining fortuitous preservation conditions with cutting-edge genomics, we map a path – narrow and winding – toward reading the genetic remnants of one of Earth’s most famous giants. It’s a quest at the very edge of plausibility. Yet, with each technical advance and each exceptionally preserved fossil, what once seemed impossible moves closer to the realm of the possible. Someday, in the not-too-distant future, a few base pairs gleaned from a T. rex might rewrite what we know about dinosaur biology, turning a tiny surviving sequence into a roaring insight from the Mesozoic past. References: (scientific support and analogous studies) Allentoft et al. (2012). Half-life of DNA in bone: DNA decay kinetics in fossils – extrapolated survival limits en.wikipedia.org . Penney et al. (2013). Study showing absence of DNA in copal/amber inclusions, undermining Jurassic Park scenario sci.news sci.news . Cano & Poinar (1993). Claim of DNA from 120–135 Myr amber weevil – initial excitement about amber as DNA preservative latimes.com latimes.com . Willerslev et al. (2022). Recovery of 2-million-year-old DNA from Greenland permafrost – current record for oldest DNA sequenced scientificamerican.com scientificamerican.com . Schweitzer et al. (2005–2012). Discovery of soft tissues and collagen in T. rex and hadrosaur bones news.ncsu.edu nature.com ; detection of possible dinosaur DNA signals via histochemistry news.ncsu.edu . Bailleul et al. (2020). Preservation of cartilage cells with nuclei in 75 Myr Hypacrosaurus – evidence of DNA and chromosomes in dinosaur cartilage news.ncsu.edu news.ncsu.edu . Saitta et al. (2019). Mechanistic studies on iron and glycation cross-linking preserving dinosaur vessels nature.com nature.com . Demarchi et al. (2016). Sequencing of 3.8 Myr ostrich eggshell proteins, stabilized by mineral binding atlasobscura.com . Various ancient DNA methodological papers: on ultra-short DNA sequencing, damage repair, and computational assembly of highly degraded genomes sci.news spaam-community.org . These underscore that techniques are rapidly improving to tackle ever older and smaller DNA fragments. |